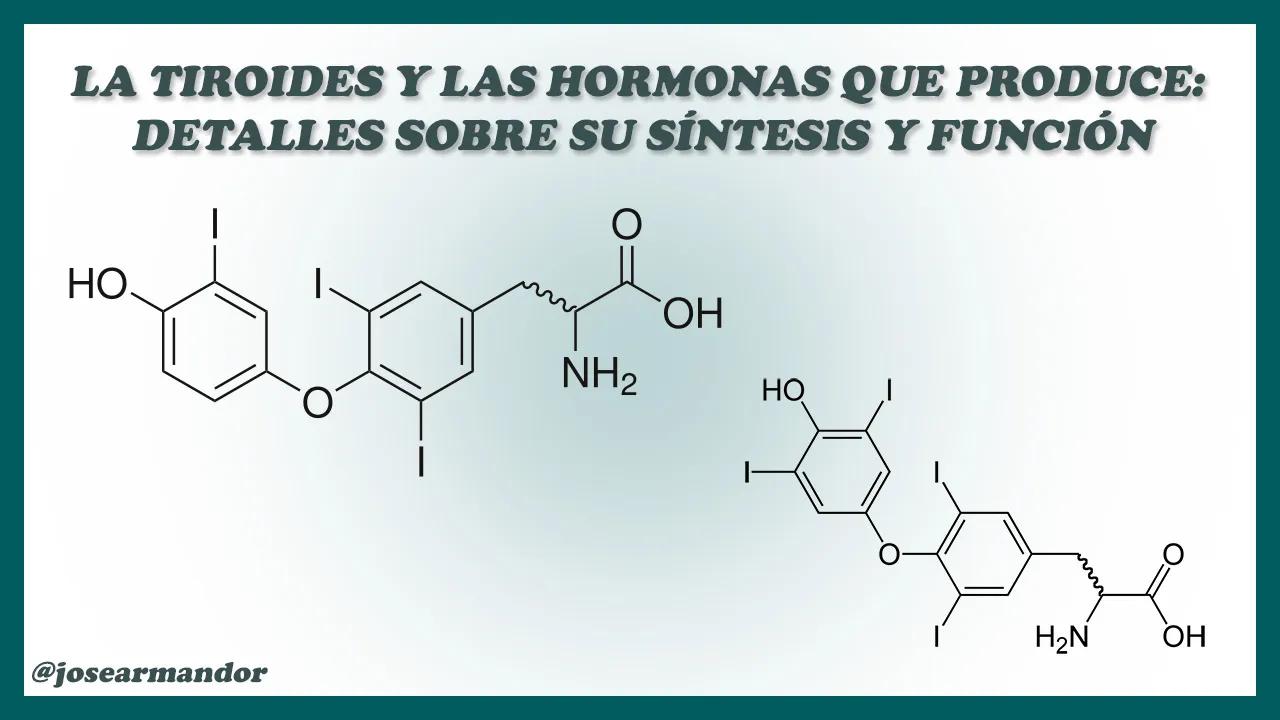
En la imagen se muestran las estructuras moleculares de la Triyodotironina y de la Tiroxina. Las imagenes de las estructuras son de dominio público bajo licencia CC0. Compilación elaborada por @josearmandor con Adobe® Photoshop® 6.
Saludos estimados compañeros de la comunidad, siguiendo en la línea de mis últimas publicaciones, acerca de la regulación hormonal en vertebrados, esta vez les traigo un trabajo de investigación donde describo los aspectos fundamentales de la anatomía de la glándula tiroides, desde los vertebrados basales hasta los humanos, adicionalmente trataré con especial detalle los procesos de síntesis de las hormonas tiroideas, así como sus principales funciones.
INTRODUCCIÓN:
La tiroides está presente en todos los vertebrados, aun cuando su forma y posición anatómica varía ampliamente en los mismos. En algunos de los grupos de vertebrados inferiores los folículos tiroideos están presentes, aunque no están organizados dentro de una glándula encapsulada y compacta. Los folículos tiroideos de la lamprea y los teleósteos tienden a diseminarse a lo largo de la aorta ventral, encontrándose frecuentemente sobre las arterias branquiales. La tiroides de algunos peces es muy móvil y puede localizarse dispersa entre la región faríngea y áreas tan alejadas como el ojo, el encéfalo, el bazo y el riñón. Contrariamente a lo mencionado en relación a ciclóstomos y teleósteos, la tiroides de los elasmobranquios es un órgano encapsulado e impar situado por debajo de la faringe. Los folículos tiroideos están ausentes en invertebrados y protocordados.
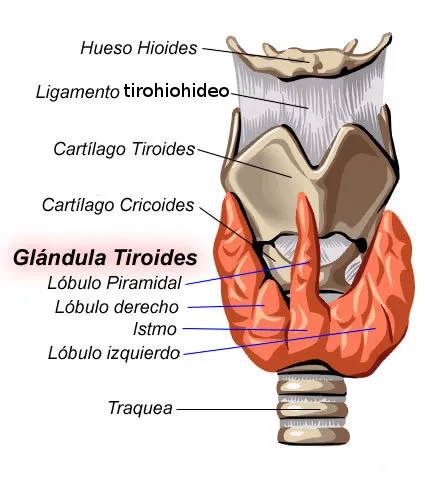
La tiroides humana se desarrolla a partir del piso de la boca embrionaria, como un divertículo medio que crece hacia abajo a manera de conducto tubular, con un extremo bifurcado que forma el istmo y parte de los lóbulos laterales, derivando el resto de estos últimos a partir de la bolsa faríngea. En ocasiones hay un lóbulo piramidal que se origina en el istmo, enfrente de la laringe. El conducto tirogloso que conecta con la boca desaparece durante el desarrollo. En el adulto, la glándula está situada a nivel del extremo inferior del cuello, frente a los anillos superiores de la tráquea, prolongándose ocasionalmente hacia el tórax por detrás del esternón (Fig. 1). El peso de la glándula tiroides en el humano adulto es de aproximadamente 25 g y consiste en un enorme número de sacos o folículos, de formas diversas desde redondeadas hasta tubulares. Su diámetro oscila entre 0,05 y 0,5 mm. Cada folículo está rodeado por una membrana basal y su pared está compuesta por una capa de células epiteliales cuboidales. Los folículos se localizan muy próximos, en una malla de tejido conectivo que contiene una red capilar extensa. Se calcula que de 4 a 6 ml de sangre por gramo de tejido perfunde la glándula normal por minuto, cantidad que se incrementa grandemente cuando la glándula se encuentra hiperactiva. En total, recibe aproximadamente el 1% del rendimiento cardíaco por minuto. Los folículos se encuentran ocupados por un material proteico semifluido, el coloide (Fig. 2). La inervación de la tiroides la efectúan fibras de los sistemas simpático y parasimpático.
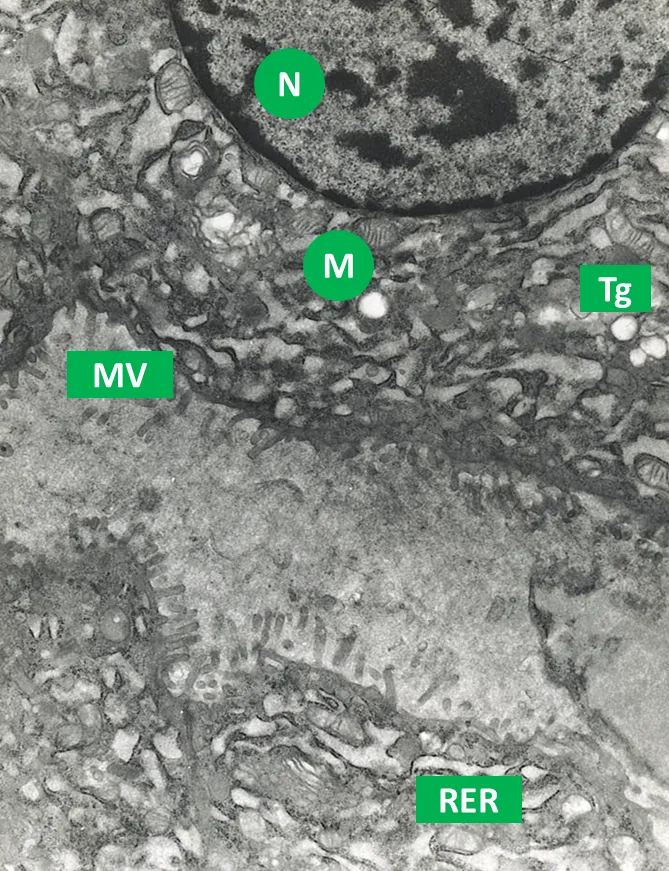
Fig. 2. Sección de un folículo tiroideo. En esta micrografía: N: Núcleo; M: Mitocondria; RER: Retículo endoplasmático rugoso; MV: Microvellosidades que limitan con el coloide; Tg: Tiroglobulina. Aumento 30.000X. Imagen obtenida en el Centro de Microscopía Electrónica de la Facultad de Ciencias de la UCV. Propiedad de los coautores: @josearmandor y Dra. Norma Ortega. Publicada bajo el consentimiento de los coautores.
LAS HORMONAS TIROIDEAS:
Las hormonas producidas por la tiroides son la Tiroxina (T4) y la Triyodotironina (T3). Ambas derivan del aminoácido tirosina, el cual yodado forma parte de la Tiroglobulina (Tg). Esta última constituye la mayor parte del coloide y posee un peso molecular de 670 kDa, con un contenido de carbohidratos del 10%. El número de aminoácidos tirosina por molécula es de alrededor de 120. La hidrólisis de la Tg produce varios derivados yodados, incluyendo la T3 y la T4. En la Fig. 3 se muestra el mecanismo por el cual se biosintetizan las hormonas tiroideas, a partir del aminoácido tirosina.

Fig. 3. Mecanismos de síntesis de las hormonas tiroideas. Propuesto por Chan y Kilby, 2000. Esquema elaborado por @josearmandor con Adobe® Photoshop® 6.
La unión del yodo a los residuos de tirosil no ocurre directamente sino que el ión debe ser primero oxidado a un estado más reactivo, siendo tal oxidación catalizada por la tiroperoxidasa, en una reacción en la cual el peróxido de hidrógeno es el aceptor de electrones. El peróxido también participa en la unión del yodo a la tirosina (Fig. 4).
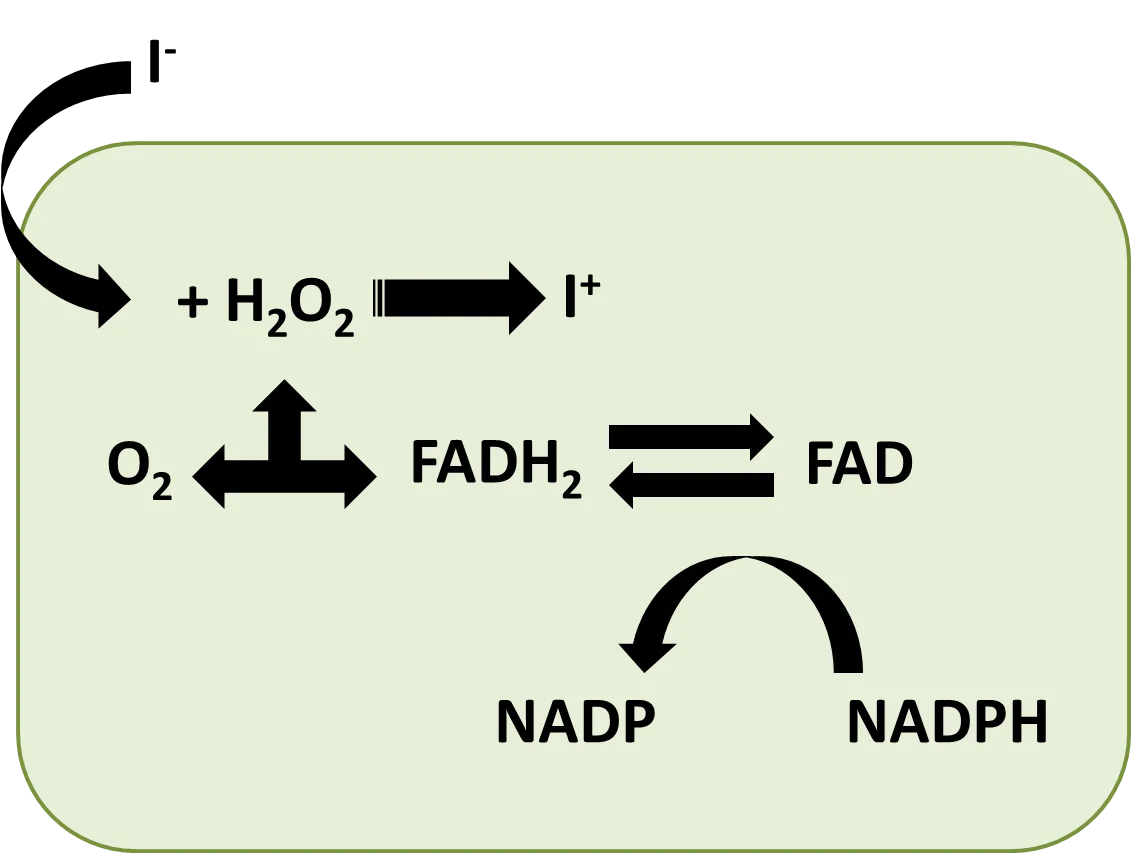
Fig. 4. Conversión del yodo en la tiroides. Propuesto por Chan y Kilby, 2000. Esquema elaborado por @josearmandor con Microsoft PowerPoint® 2010.
La tiroides concentra el yodo a partir de la sangre, mediante un transporte activo en contra del gradiente químico, y también de uno eléctrico, puesto que las células foliculares mantienen una diferencia de potencial de -40 a -50 mV a través de la membrana plasmástica. El ATP es requerido a objeto de que la concentración pueda tener lugar, estando probablemente involucrada en el mencionado mecanismo una ATPasa N/K dependiente.
La Tg alcanza su tamaño total dentro de la cisterna del retículo endoplasmático de la célula folicular (Fig. 2) y de allí es transportada hacia el aparato de Golgi (en dirección apical), siendo adicionada con unidades de carbohidrato. Las unidades centrales de oligosacáridos (conteniendo glucosa, manosa y N-acetilglucosamina) son transferidas desde el transportador lipídico a la tiroglobulina, ya en la cisterna del retículo endoplasmático, en tanto que los azúcares periféricos (galactosa y ácido siálico) parecen ser añadidos en el aparato de Golgi.

La última parte del transporte intracelular de la Tg tiene lugar en vesículas apicales exocíticas. Estas últimas descargan su contenido a la luz folicular. Además de la Tg, en las reacciones de yodinación intervienen: yodo, una peroxidasa y el peróxido de hidrógeno. La peroxidasa de la tiroides es una glicoproteína, cuyo grupo prostético es un componente hemo y se localiza firmemente unido a la superficie externa de la membrana plasmática, en forma integral. La tiroides tiene capacidad de generar peróxido de hidrógeno, habiéndose propuesto varios sistemas enzimáticos como responsables de tal producción.

La yodinación de la Tg probablemente tiene lugar en la luz folicular en asociación con la membrana apical de la célula folicular. La peroxidasa, presente en la membrana plasmática apical, es importante para todas las etapas en la síntesis de las hormonas tiroideas, incluyendo no sólo la oxidación del yoduro sino también su unión al tirosil en la Tg y la formación intramolecular de yodotironinas por acoplamiento de los residuos yodados de tirosil. La liberación de las hormonas tiroideas a la sangre se inicia con la captura del coloide por la célula folicular a través de un proceso de endocitosis, existiendo el modo micropinocítico y el macropinocítico. Dentro de la célula folicular, los endosomas se fusionan con los lisosomas y las hormonas tiroideas son liberadas de la Tg por acción de enzimas proteolíticas. No se conoce el mecanismo por el cual las hormonas tiroideas son transferidas desde el compartimiento lisosomal hasta la membrana plasmática basal y luego vertidas a los capilares.
DATOS INTERESANTES SOBRE LA T3 Y LA T4:
TRIYODOTIRONINA (T3):
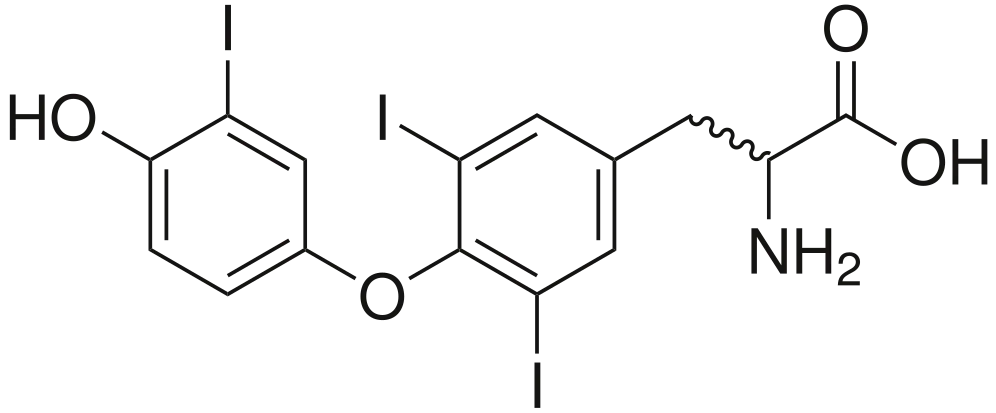
• Representa únicamente el 5% del total de las hormonas tiroideas circulantes.
• Un tercio de la T3 presente en la sangre proviene de la conversión periférica de la T4 a T3, en tanto que dos tercios de ella derivan de los folículos tiroideos.
• La T3 es más activa que la T4 (la relación T3:T4 es 1:4) y se une débilmente a la globulina transportadora de hormonas tiroideas (TBG).
• La T3 tiene una vida media de aproximadamente 1 día, la cual resulta más corta que la de la T4.
• El 99,70% de la hormona está unida, mientras que el 0,30% está libre.
TIROXINA (T4):

• Representa el 95% del total de las hormonas tiroideas circulantes.
• Está unida fuertemente a las proteínas plasmáticas TBG (60-75%), TBPA (30-15%) y albúmina (10-5%).
• La afinidad de la TBG por la T4 es de 2 a 6 veces mayor que por la T3.
• La vida media de la T4 es de aproximadamente una semana.
• El 99,96% de la T4 circulante está unida.
FUNCIÓN Y REGULACIÓN:
Las hormonas tiroideas (T3 y T4) estimulan el crecimiento y son factores fundamentales en la condición homeoterma de aves y mamíferos. Las hormonas tiroideas poseen una acción que es sinérgica con la de la hormona de crecimiento, en el sentido de que mientras esta última tiene efectos sólo en la estimulación del crecimiento longitudinal, las primeras inducen la maduración. Tales efectos se llevan a cabo por la vía de modificar a nivel nuclear cualitativamente y cuantitativamente la síntesis proteica. La T3 y en menor grado la T4, se unen a las proteínas nucleares no histónicas e inician el proceso de transcripción génica y la síntesis de ARNm.
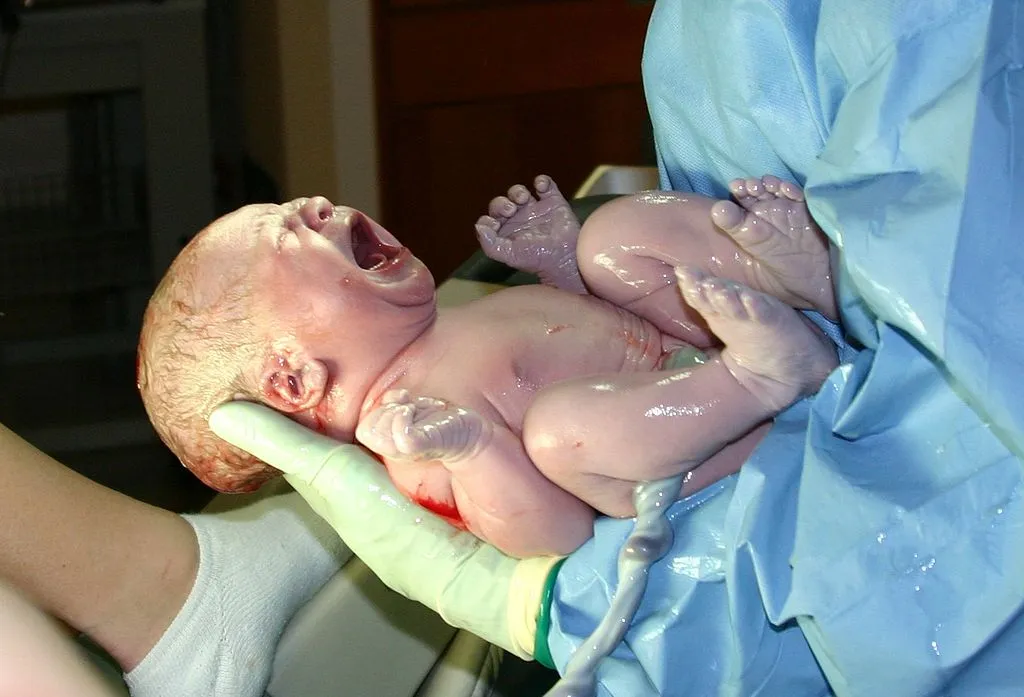
Está bien establecido que el status tiroideo de los neonatos y niños tiene a largo plazo un significativo impacto sobre su comportamiento, habilidad locomotora, lenguaje, oído y cognición. Retrasos en la restauración del status tiroideo en el neonato pueden conducir a un daño irreversible. Asimismo, el desarrollo de diferentes áreas del sistema nervioso central se ha asociado con el tiempo y la duración de deficiencias en hormonas tiroideas, lo que sugiere que hay períodos críticos durante los cuales varias partes del cerebro son sensibles al suplemento de hormonas tiroideas. Imagen de dominio público bajo licencia CC0.
Las hormonas tiroideas son únicas en relación a su habilidad para estimular la termogénesis basal. Para explicar el mecanismo por el cual las hormonas tiroideas estimulan la termogénesis, se ha sugerido que a nivel molecular el mecanismo principal es el control de genes específicos a nivel transcripcional. Actualmente, el conocimiento de los mecanismos subyacentes de la termogénesis y su control involucran directa o indirectamente al aparato de transducción de la energía en las mitocondrias. Desde hace más de 50 años, se ha sugerido que las hormonas tiroideas incrementan la tasa metabólica al desacoplar el transporte electrónico asociado a la síntesis de ATP. Esta hipótesis fue desechada ya que se pensó que no era fisiológicamente relevante: los efectos vistos in vitro no eran observados in vivo. Sin embargo, últimamente la hipótesis del desacoplamiento ha recibido nuevo apoyo por las siguientes razones:

En las mitocondrias, el consumo de oxígeno no está perfectamente acoplado a la síntesis de ATP, ya que parte del gradiente electroquímico protónico se disipa como calor. Dos mecanismos podrían explicar ese acoplamiento imperfecto: reentrada pasiva de protones a la matriz (fuga de protones) y una falla de las bombas de protones, consistente con una salida reducida de protones, a pesar de presentarse una similar tasa de transporte electrónico. Actualmente, está bien establecido que la fuga de protones es una propiedad general de la membrana mitocondrial interna. Imagen de dominio público bajo licencia CC0.
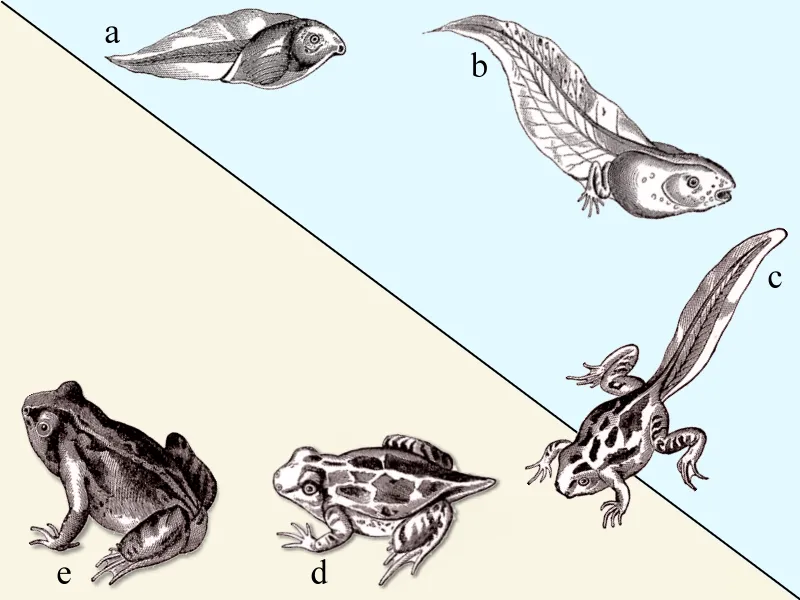
Se descubrió que proteínas desacoplantes están presentes en casi todos los tejidos, y debido a su involucramiento en los mecanismos que operan en la termogénesis celular, ellas son candidatas obvias como mediadoras de la termogénesis tiroidea. En los anfibios, las hormonas tiroideas estimulan la metamorfosis, proceso por medio del cual la forma acuática del renacuajo se transforma en la terrestre del anfibio. Precisamente antes del inicio del clímax metamórfico, es que se incrementa grandemente la secreción de esas hormonas. Imagen de dominio público bajo licencia CC0.
Bibliografía consultada:
• Chan, S. & Kilby, M.D. (2000). Thyroid hormone and central nervous system development. J. Endocrinol. 165: 1-8.
• Dickey, R.A.; Wartofsky, L. & Feld, S. (2005). Optimal thyrotrophin level: Normal ranges and reference intervals are not equivalent. Thyroid. 15: 1035-1039.
• Hoffmann, G. & Dietzel, I.D. (2004). Thyroid hormone regulates excitability in central neurons from postnatal rat. Neuroscience. 125: 369-379.
• Lanni, A.; Moreno, M.; Lombardi, A. & Glglia, F. (2003). Thyroid hormone and uncoupling proteins. FEBS Letters. 543: 5-10.
DESCUBRE OTROS ARTÍCULOS INTERESANTES VISITANDO LA ETIQUETA #stem-espanol.

Imagen elaborada por @josearmandor para la comunidad #stem-espanol. La imagen utilizada en el fondo es de dominio público.
